- ... `Zerox'2.1
- From the album Dirk Wears White Sox by Adam and the Ants ©1979 Do-It Records.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... position.2.2
- The major
extension that produced the neo-Darwinist position was the synthesis
of Darwin's original theory with Mendelian genetics.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... forms.2.3
- Darwin did not claim his theory
accounted for the origin of life. However, more recently, a
number of theories have been proposed which put the original
transition from chemistry to biology in an evolutionary context
(see, for example, [Eigen 71],
[Maynard Smith & Szathmáry 95]).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
emphasis).2.4
- The enclosed quotation is from [Oyama 85].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Species2.5
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Species2.6
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Species2.7
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
- The ideas in this section are partially based upon a
talk given by Richard Dawkins at the `Digital Biota 2' conference in
Cambridge, England on 10th September 1998.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... impossible.2.8
- In fact, it is still not
entirely clear whether the existence of species is a consequence of
sexual reproduction ([Maynard Smith & Szathmáry 95], Chapter
9), but this does not affect the argument being made in this
section.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
literature,2.9
- For discussion of this, see, for example,
[Margulis 91], [Daida et al. 96].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... prokaryotic2.10
- Present-day
bacteria are examples of prokaryotic cells.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... eukaryotic2.11
- All present-day animal, plant and fungi
cells are eukaryotic. They are far more complex than prokaryotic
cells, housing a wide array of specialised structures.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... flux.2.12
- Although the extent to which
this is true varies greatly between different organisms, and
between different parts within a single organism
[Maynard Smith 86] (p.2).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... systems'2.13
- They invented the word
`autopoiesis' from the Greek
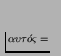 self, and
self, and
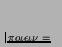 to produce.
to produce.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... is:2.14
- Unfortunately the
definition, at least in its English translation, is somewhat opaque.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... changed.''2.15
- Quoted from
[Kelly 94] (p.447).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Earth.2.16
- Here I am only talking about models that assume a
terrestrial origin of life.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... conditions)2.17
- One necessary
condition is that the self-maintaining organisations were enclosed
in some form of compartment, to enable natural selection to act
between the compartments [Szathmáry & Demeter 87].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
speciation,2.18
- That is, speciation occuring as the result of the
restriction (or total absence) of gene flow between two populations
caused by some sort of physical barrier (e.g. a sea or mountain
range). See, for example, [Maynard Smith 89].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... theory.2.19
- In the terminology of evolutionary
algorithms, Wright's theory is a mechanism whereby an evolving
population can avoid getting stuck in local (sub-optimal) fitness
peaks. It involves the population being divided into a large number
of small, partially isolated demes. Individuals in one deme may move
to a higher fitness peak by chance (i.e. genetic drift), and
subsequently spread throughout the whole population. See, for
example, [Wright 31] and [Wright 82]; and also
[Coyne et al. 97].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Thompson 17].2.20
- Quoted from
[Nitecki 88] (p.14).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Darwin 72].2.21
- Quoted from
[Gould 89] (p.257).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... complexity.2.22
- McShea also mentions a third possible
dichotomy: differentiation versus configuration. The four types of
complexity mentioned above are differentiational. Configurational
complexity, on the other hand, is ``irregularity of arrangement of
parts and interactions, independent of their differentiation''
[McShea 96] (p.480). McShea claims that
configurational complexity has received little attention in the
biological literature, so he does not consider it further in his work.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Bronowski 73].2.23
- Although Bronowski used the term in a
somewhat broader context.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... applies.2.24
- Note that I am not talking
about the distinction between gradualism and punctuated equilibria
here; this view of evolution does allow for evolution in the periods
between major transitions. However, the view holds that evolution
during these periods will not necessarily be in the direction of
increased complexity.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Gould 89].2.25
- Although Gould's view is also
connected to the concept of punctuated equilibria, which is not
specifically related to transitions between units of selection.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... next).2.26
- Salthe, in considering
the hierarchical nature of life, argues that there is a clear
distinction between the ecological hierarchy and the genealogical
(evolutionary) hierarchy
[Salthe 85]. The confusion in
terminology under discussion here can also be seen as a confusion
between entities in these two distinct (but related) hierarchies.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... process.2.27
- That
is to say, S-lineages are the fundamental types of things
which act as units of selection. McMullin is not arguing here about
whether it is more appropriate to look at evolution from the point
of view of the gene or the organism, for example, but claims that
``whichever of these viewpoints may be adopted, there will be a
crucial distinction between actors and S-lineages, with only the
latter being properly regarded as units of selection, or entities
for whose benefit (Darwinian) adaptations may be said to exist''
[McMullin 95] (p.166, original emphasis).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Karakotsios3.1
- Quoted from [Kelly 94] (p.451).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Langton 86]),3.2
- Although
others had used the term in earlier publications, such as
[Overton 82] (in which it appears in the title without
definition in the text, and in fact, by comparing the contents of
the main text with that of the editor's note at the beginning, it
appears that the title (including the term `artificial life') was
chosen by the editor, Edmund C. Berkeley, rather than by Overton
himself).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... subject3.3
- Although some would
argue over the extent to which it is a coherent subject. See
Sections 3.1.1-3.1.3.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... purposes,3.4
- For example, see Steven
Rooke's online portfolio at
http://www.concentric.net/~Srooke/ and the list of genetic
art-related sites at
http://gracco.irmkant.rm.cnr.it/luigi/alg_art.htm.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... p.372).3.5
- The concern with
self-replication has also been a preoccupation for other artificial
life researchers from von Neumann [von Neumann 66] to the present
day, and can be related to the work of biologists such as Muller and
Dawkins, discussed in Section 2.2.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... terms,3.6
- This, of course, is a big
assumption. Those who maintain that life is of necessity a
biochemical phenomenon would argue that concepts such as autopoiesis
do not capture all of the relevant features of life. However,
this eventually boils down to a linguistic debate, and scientific
progress can only be made when we do adopt precise definitions, even
if these are not universally agreed upon.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... example).3.7
- But note that artificial life
does not restrict itself to using digital computers as the medium of
implementation.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... axioms)3.8
- David Marr's
[Marr 82] analysis of complex systems on three levels (the
computational theory level, the representation and algorithm level, and
the hardware implementation level), and his insistence that the
theoretical distinction between these levels should be recognised when
devising a model--especially the distinction between what is being
computed (level 1) and how (level 2)--is also relevant for the
formulation of scientific artificial life models.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... machines.3.9
- Von Neumann had
difficulties in defining precisely what the term `complicated'
meant.
He said ``I am not thinking about how
involved the object is, but how involved its purposive operations
are. In this sense, an object is of the highest degree of complexity
if it can do very difficult and involved things.''
[von Neumann 66].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... above.3.10
- The general constructive
machine A of this design is often referred to as a `universal
constructor'. However, this term should be used with caution; from
the above description of the architecture it is clear that A
can build any machine X that can be described upon a
tape
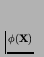 . For cellular automaton models it can be
proved that there are some configurations that the universal
constructor cannot build (e.g. [Moore 62],
[Myhill 63]). These are referred to as `Garden
of Eden' configurations, as the only way they may exist is if they
are programmed in as the initial state of the space at time zero.
. For cellular automaton models it can be
proved that there are some configurations that the universal
constructor cannot build (e.g. [Moore 62],
[Myhill 63]). These are referred to as `Garden
of Eden' configurations, as the only way they may exist is if they
are programmed in as the initial state of the space at time zero.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[von Neumann 66].3.11
- Quoted from
[Pattee 88] (p.69).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... self-reproduction3.12
- `Trivial' self-reproduction, at
least in the sense being used by Langton, occurs
when reproduction of a particular sort of configuration happens
purely due to the rules of the system rather than to anything
explicitly encoded in the configuration itself. For example, a CA
with a transition rule such as ``if this cell is empty (in the
quiescent state) and one of its neighbouring cells is in state
 ,
then change the state of this cell to '' is an
environment in which the state
trivially
self-replicates. I suspect that Langton's work was the seed for
the preoccupation of many artificial life researchers over the past
decade with the `problem of trivial self-reproduction'. However, I believe
that this is not the most relevant distinction with respect to
evolvability, as I will argue in Chapter 7.
,
then change the state of this cell to '' is an
environment in which the state
trivially
self-replicates. I suspect that Langton's work was the seed for
the preoccupation of many artificial life researchers over the past
decade with the `problem of trivial self-reproduction'. However, I believe
that this is not the most relevant distinction with respect to
evolvability, as I will argue in Chapter 7.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... 1990s.3.13
- Although Eigen
and Schuster's work in the 1970s should also be mentioned. In the
context of prebiotic evolution, they discussed ways in
which the amount of selectively maintainable information in a system
could be increased beyond the capacity of an individual replicator
(see, for example, [Eigen & Schuster 77]). They introduced the
`hypercycle' (a cyclically catalytic group of replicators) as an
functional organisation which could achieve this.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
`Coreworld'3.14
- Specific configurations of Coreworld were named
Venus I, Venus II, and Luna.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Dewdney 84].3.15
- The
ancestry of this approach can be traced back even earlier
(e.g. [Bratley & Millo 72], [Burger et al. 80]),
although these examples were presented in a fairly informal manner.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... universality.3.16
- A proof that the language has
this property is presented in [Maley 94].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
template-driven3.17
- With the template-driven branching scheme, a
branching instruction is followed by a template (a sequence of
bits). The operating system will search for the nearest matching
template in the rest of the code and move the instruction pointer to
that position.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... problem,3.18
- That is, some programs might never terminate.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... set3.19
- Examples include issuing a jmp
instruction with a template pattern for which no match can be found,
and attempting to write to a memory address for which the program
does not have write access.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... sin.''3.20
- From an online report on Tierra,
available at http://www.hip.atr.co.jp/~ray/tierra/netreport/
netreport.html#Philosophy.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
symbiogenesis3.21
- According to Barricelli, the symbiogenesis
theory claims that if genes are to evolve into ``relatively higher
forms of life'', they must only be able to reproduce through a
symbiotic relationship with other genes
[Barricelli 57] (p.145). A group of genes
that collectively reproduces in this way (a `symbioorganism') can be
considered a special case of a hypercycle
[Eigen & Schuster 77], in which each component in the group is
absolutely required for the reproduction of the next
component, rather than just acting as a catalyst.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... community,3.22
- His papers
contained many ideas on subjects which have since become popular
research areas, such as the use of real DNA for performing
computations ([Barricelli 63] pp.121-122).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Bedau 98a]).3.23
- But see [Mayer & Rasmussen 98] for
a recent study in which emergent hierarchical structure has
been successfully simulated.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[McMullin & Varela 97].3.24
- Interestingly, Barricelli also
predicted that his symbioorganisms would need to develop means of
controlling their local environment (such as a membrane) if they
were to evolve past a certain level of complexity
[Barricelli 63] (pp.122-124).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
coexisted.3.25
- Another is Lindgren's model of a population of
individuals playing a variation of the iterated Prisoner's Dilemma
[Lindgren 91].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Buss.3.26
- Rasmussen et al. also discuss the
relationship between these and other universal formalisms
[Rasmussen et al. 91] (pp.243-244).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
structures'';3.27
- This is similar to von Neumann's conclusion
that ``complication is degenerative below a certain minimum level''
[von Neumann 49] (p.482).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... principles.3.28
- More precisely, there may be some
general principles concerning parasitism that Tierra shares with
Nature, but the point is that no-one has explicitly stated what
these might be, and Tierra was therefore not designed to be a
particularly good test of any such principles.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... sort.3.29
- Maynard Smith and
Szathmáry discuss ways in which the evolutionary potential
of hypercycles can be enhanced through compartmentation, and suggest
that this might eventually lead to the evolution of a genetic system
[Maynard Smith & Szathmáry 95].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Cosmos,4.1
- `Cosmos' is an
acronym for COmpetitive Self-replicating Multicellular Organisms in
Software.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... complexity4.2
- Primarily, hierarchical object
complexity (see Section 2.3.2). In other words, many
of the design features of Cosmos were intended to promote the
evolution of multicellular organisms from unicellular ones.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... squares.4.3
- The
system has been designed to deal with arbitrary n-dimensional
environments, but the current implementation requires some minor
revisions to allow this.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... execute.4.4
- As I am usually
referring to the contents of the Genome, rather than to the
structure itself, when I use the term `genome', I will use the
standard typeface from now on.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
instruction),4.5
- See Section A.2 for an
explanation of the instruction set.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... genome4.6
- Or on
eligible InfoStrings in the Received Message Store. See
Section 4.3.7 for details.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
string.4.7
- The search begins at the current Read position on the
genome, and proceeds outwards in both directions
simultaneously.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
genome,4.8
- Or on eligible InfoStrings in the
Received Message Store. See Section 4.3.7 for details.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... tokens,4.9
- In this situation, the choice of which cells
to kill is actually stochastic, with the level of a cell's
Energy Token Store determining the probability of its being killed.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... area,4.10
- The only restriction is that there is a maximum
length to which these strings are allowed to grow, defined by the
global parameter info_string_size_limit. This is to prevent
the situation in which a
program evolves which gets stuck in an infinite loop writing to the
Nucleus Working Memory, eventually using up all of the memory in the
system.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...)4.11
- The function of
these latter two structures is explained in Section 4.3.7.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... it.4.12
- The type of
the cell's genome cannot be directly altered, and is passed on to
children when the cell splits or divides. However, it is subject to
mutation like any other part of the cell (see
Section 4.5.7). Therefore, it is possible
for organisms with different genome types to emerge in the
system.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...Organism4.13
- A
capital `O' is used here to emphasise that we are talking about the
specific implementation details. However, as the Organism class
encapsulates the functionality of an organism, the two terms can be
used interchangeably. Therefore, in the rest of the document I shall
just use the term organism (with a small `o'). The same applies for
cells and the Cell class.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
REPLiCa,4.14
- `REPLiCa' is an acronym for Robust Evolvable
Programming Language for Cosmos.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... promoter.4.15
- Of course, when
programs are evolving, especially when we are considering parallel
programs, there may be more than one promoter in the Promoter Store at one time. However, here we are describing how a human might
design a program that performs a jump--evolution would probably go
about designing a program in a very different way.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... amount.4.16
- To be precise, the magnitude of the
increment is kIp, where I is the current intensity, and k and
p are constants defined by the global parameters
envinfostring_decay_constant and
envinfostring_decay_power respectively.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... limits.4.17
- To be precise, the flaw rate can change by
plus or minus n parts per thousand, where n is determined by the
parameter flaw_period_max_change_per_thou.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... genome.4.18
- Tierra
features both mutations and flaws (although the
mechanisms for flaws is somewhat different) but in
subsequent work by Chris Adami and Titus Brown with their Avida
system the authors suggested that flaws played only a minor role
in evolution compared to mutations [Adami & Brown 94].
Informal observations from preliminary runs of Cosmos suggested
that flaws in the execution of instructions significantly increase the
rate at which useful mutants are produced.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
rate5.1
- Precisely, what matters is the rate at which
selectively significant errors occur, i.e. errors which
exclude the offspring from that particular S-lineage.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
rate,5.2
- Actually, we looked at the inverse of flaw rate, the flaw
period. This is the expected number of successful instruction
executions in a program between successive flaws.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[5.3
- Note we are looking at genotypes here,
i.e. groups of identical programs, rather than individual programs.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Bedau & Brown 97]5.4
- The rewording at
the indicated places in this extract is to make it consistent with
later changes in the terminology that Bedau et al. use to describe
these techniques.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
- Again, this formula has been changed from the
original text to make the notation more consistent. In the original
paper, tex2html_wrap_inline$A_new(t)$ is defined as displaymathA_new(t) =
1D(t)_i,a_0a_i(t)a_1a_i(t)
(i.e. as above, but divided by tex2html_wrap_inline$D(t)$). This is what
will be referred to here as Mean New Activity,
tex2html_wrap_inline$A_new(t)$.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Bedau et al. 98]5.5
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... unicellular.)5.6
- Therefore, at any given time, the number of
cells in the neutral shadow is always the same as in the standard
run, although the number of organisms may be lower if the standard
run contains some multicellular (parallel) programs.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
bits.5.7
- There were in fact a handful (7) of programs of
different lengths, but these are not recorded in
Figure 5.2, because of the data pruning methods
described in Section 5.1.1.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... length.5.8
- Again, the data pruning techniques
discussed in Section 5.1.1 mean that
information is not displayed in these graphs for genotypes that are
represented by only a very few individual programs. Such
individuals typically account for about 1% of the total
population.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
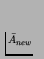 .5.9
.5.9
- As
explained earlier, Bedau calls this measure simply the new activity,
Anew, rather than mean new activity. The name has been changed
here to make it consistent with the naming of other measures.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... bounded5.10
- The definition of
(un)boundedness given in [Bedau et al. 98] is: The function f(t) is unbounded
iff
where sup(.) is the supremum function.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... positive5.11
- Similarly, the
definition of a positive function is given in
[Bedau et al. 98] as follows: The function f(t) is positive iff
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Chance6.1
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Chance6.2
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Chance6.3
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
- This section is based upon a previously published paper, [Taylor & Hallam 98].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ....6.4
- These
experiments were run on slower machines than most of the others,
which is why the size of the runs (i.e. the maximum number of cells
allowed in the population and the run duration) is smaller
than most of the others. The other differences in parameter values
came about because these runs were actually conducted before most of
the others, and the default values of a few of the parameters were
changed in the intervening time. The differences are only minor, and
it is not expected that they affect the applicability of the results
to the other experiments reported in this chapter.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
compared.6.5
- In the following, the pairs of run results displayed in
Figure 6.1 and in
Figure 6.8 were generally chosen because they
illustrate noticeably different results.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... allocated.6.6
- Recall
from Chapter 4 that under this scheme, the
parameter
number_of_energy_tokens_per_ grid_pos_per_sweep
determines the mean number of energy tokens that should be
distributed to each grid position.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
instruction6.7
- Or sometimes a migrate instruction,
which is equivalent to move for a single-celled program.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
readable.6.8
- In contrast, the mean number of faithful reproductions of
programs of the ancestor genotype 348AAAA were similar in the two
sets of experiments. In the present experiment, the figure was 1.077
(standard deviation 0.045), and in the standard model re-runs, it
was 0.993 (standard deviation 0.037).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
energy.6.9
- This section was included for the practical reason
that when a run is first inoculated, several hundred time slices
pass before the females emit their first messages. During this time,
collecting energy is the only useful task that the males can do.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... speciation6.10
- I am using the
term `speciation' very loosely here. For a start, as the programs
are reproducing asexually, it is debatable whether the concept of
species is applicable at all. Even if we do allow the term to be
used, it might be argued that the
programs in the different length groups are fundamentally very
similar, and are more appropriately viewed as different varieties of
the same species. However, remember that they did not only differ in
their lengths, but also sometimes in their ability to move, and by
the amount of energy they collected.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... issue,6.11
- They were not designed with this in mind. A proper
test would need to be based upon an explicit set of assumptions, and
would almost certainly require sexually reproducing programs.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... environment.6.12
- This
theory, which he named `cannibalistic altruism', was discussed
during a talk by Wolpert at the Royal Museum of Scotland,
Edinburgh, on 20 February 1997.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
reserves),6.13
- But note that in this case the prey organisms
would only be a resource of energy, not of matter. This distinction
is discussed in more detail in the next chapter (e.g. Section 7.1.4), along with possible evolutionary
consequences arising from it.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... model).7.1
- Ray himself recognises these difficulties, but
is more optimistic that they can be overcome [Ray 91] (p.399).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
(p.36).7.2
- Although he also stresses that ``some epistemic
principles must restrict physics-as-it-could-be if it is to be any
more than computer games'' [Pattee 95a].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
organisms).7.3
- By `environment' I mean the shared space in which
(at least some aspects of) all of the organisms exist, which I will
refer to as the `arena of competition' (see
Section 7.2.3), together with the universal `laws
of physics' of this space which determine how entities within it act
and interact.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... information.7.4
- For example, a virus requires
information contained in its host's genome in order to
reproduce. This information is more than the matter from which the
host's DNA is constructed; it involves a particular ordering of
matter.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
matter.7.5
- This is also the case in von Neumann's cellular
automata models. In contrast, structures in his kinematic model, as
in models proposed by various others (such as Myhill, and Holland's
 -Universes), are constructed from atomic units of matter (see
Chapter 3).
-Universes), are constructed from atomic units of matter (see
Chapter 3).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Rosen 59]).7.6
- Although Löfgren has shown
that complete self-reproductive functions can exist through
axiomatisation [Löfgren 68].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... division).7.7
- The
important distinction between these two types of system in the
present context is that logical systems manipulate states, whereas
material systems manipulate matter. Reproduction in material
systems, in contrast to logical systems, therefore requires an object
to collect the `raw materials' to build a copy of itself.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
distinction.7.8
- Any attempt to classify a reproducer as
trivial or non-trivial according to the explicit-implicit
distinction is bound to be somewhat arbitrary, because we are
generally considering examples of second-level reproduction rather
than complete self-reproduction (Section 7.2.1). Therefore, the
surroundings will always play some role in bringing about the
reproduction of the object in question. There is considerable irony
in much of the recent work relating to this issue. For example,
Langton not only ignores von Neumann's actual solution to the
`problem' of trivial reproduction, but furthermore says that ``von
Neumann's work suggests an appropriate criterion ... : the
configuration must treat its stored information in ... two different
manners ... : interpreted, as instructions to be executed ...,
and uninterpreted, as data to be copied''
[Langton 84] (p.137). As far as von Neumann's
actual analysis of the subject is concerned, this distinction
between interpreted and uninterpreted is only important insofar as
it gives the reproducing automaton the capacity to support inheritable
mutations, potentially leading to the evolution of more complicated
and more efficient machines.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...C,7.9
- This is not an inherent property of the architecture
per se, but von Neumann's analysis of evolvability did assume
a `compositional' structure in the language of the tape descriptions
(see Section 3.2.1).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... (p.574).7.10
- The passage to which I
am referring is ambiguous. Ibáńez and colleagues talk about
``the possibility of using our self-inspection based reproductive
scheme as a basis for artificial evolution. This attempt is not new;
it has quite successfully been applied in other environments, and
the most paradigmatic of them is probably Tierra''
[Ibáńez et al. 95] (p.574). I take this to imply that they
also regard Tierra as an example of a self-inspection based scheme.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
design.7.11
- Indeed, for organisms in any kind of evolving
system, the notion of a phenotype fundamentally involves behaviour,
in the form of interaction with the (biotic and abiotic)
environment.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Replication.7.12
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
- Illustration © Genentech,
Inc. (http://www.gene.com/ae/AB/GG/). Reprinted with
permission.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... cell.7.13
- For
eukaryotic cells we would have to include the DNA contained in
mitochondria and, in the case of plants, chloroplasts, as well as
that contained in the nucleus. There is some
evidence that particular features of cells can occasionally be
inherited without apparently requiring a change in DNA sequence
(e.g. abnormal patterns of cilia on the surface of particular
protozoans), but such exceptions are very infrequent
[Maynard Smith 86] (pp.24-25).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... genome.7.14
- To be
precise, the initial interpretation machinery is derived from
information contained in the parent cell's genome, in the
same way as in von Neumann's self-reproducing automata.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...A.7.15
- Note however that the laws of physics and chemistry still
play a vital role in the construction of this machinery. For
example, the genome only encodes information about the linear
sequence of amino acids in any protein that it can construct. As the
ribosomes build the protein based upon this information, the growing
protein folds into a three-dimensional shape due to attractive and
repulsive forces between its subsections. Thus the primary (linear)
structure of the protein is encoded in the genome, but its secondary
and higher structure is determined by the laws of physics (and by
the manner in which it is constructed).
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
ingredient'.7.16
- His solution was to require that elements could
only reproduce in symbiotic association with other elements (see
Section 3.2.2). While this may indeed be an
important aspect of the `missing ingredient', it is extremely
doubtful that it is the only important aspect.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... GA.7.17
- The same applies to similar artificial
life platforms with two-dimensional environments, such as Avida
[Adami & Brown 94].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
addresses.7.18
- Recall that the allocation of memory for
reproduction is performed by the external operating system.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Goals7.19
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Goals7.20
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... Goals7.21
-
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
- The phrase `Beyond Digital Naturalism' is borrowed from
[Fontana et al. 94].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... addressed,7.22
- Even in the
situation where some high-level `emergent' phenomenon is under
investigation, and the model is expressed in terms of low-level
entities and interactions, it may still be viewed as a potential
explanation in the sense that it could demonstrate that no
additional entities or interactions are required to produce the
phenomenon. David Chalmers refers to such accounts as
`mystery removing' explanations [Chalmers 96].
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... p.115).7.23
- Note that
this description closely resembles Löfgren's definition of
self-reproduction given in Section 7.2.1, if we substitute
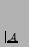 in those definitions for
in those definitions for
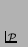 above, d for
above, d for
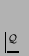 ,
and
,
and
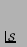 for
for
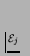 .
Sticking to
Waddington's labels, the difference is that Löfgren's definition
results in
being reproduced, whereas Waddington's results
in
being reproduced. If we assume that
is the
only aspect of
that affects the reproduction process, then
the end result is the same.
.
Sticking to
Waddington's labels, the difference is that Löfgren's definition
results in
being reproduced, whereas Waddington's results
in
being reproduced. If we assume that
is the
only aspect of
that affects the reproduction process, then
the end result is the same.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ... (p.120).7.24
- In the
original paper, the final word of this paragraph appears as
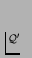 s
rather than
s
rather than
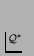 s. This is fairly clearly a typographical error.
s. This is fairly clearly a typographical error.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...[Holland 76],7.25
- Holland refers to strong and weak bonds in
his model, but the weak bond effectively denotes the absence of a
bond in the normal sense of the word.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
- ...
program.9.1
- Actiview is a program developed by
Emile Snyder and Mark Bedau at Reed College in the USA, to produce
various summary statistics and graphs depicting the evolutionary
activity of the run. These are described in
Section 5.1.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.