An evolutionary system must therefore have an arena of competition of some description, although there are few restrictions on the particular form it should take. All that is required is that it introduces the concept of (one or more) resource(s), each of which is: (a) a vital commodity to individuals in the population; (b) of limited availability; and (c) that individuals can compete for (at either a global or local level). At the lowest level, each of these resources can usually be interpreted as energy, space, matter or information (see Section 7.1.4).
An issue that arises when considering different evolutionary systems is the extent to which individuals are embedded in this arena of competition. In von Neumann's cellular automata design, individuals are fully embedded--there is no `hidden' state information (i.e. information which is not embedded in the cellular space itself). If one believes in materialism, the same can be said of the biosphere. At the other extreme, individuals in a genetic algorithm (GA) have minimal embeddedness--the arena of competition merely contains place holders for the chromosomes, and the restriction is generally on the number of individuals, regardless of their size (although most GAs have constant-size chromosomes anyway). These two extremes, together with intermediate situations arising in Cosmos and Tierra, are depicted in Figure 7.3. Note that individuals in Cosmos are not really embedded in the arena of competition at all; the two-dimensional environment only holds pointers to the cells, in much the same way as in a GA.7.17 In Tierra, a program's instructions are embedded in the arena, although each program still has some additional state information.
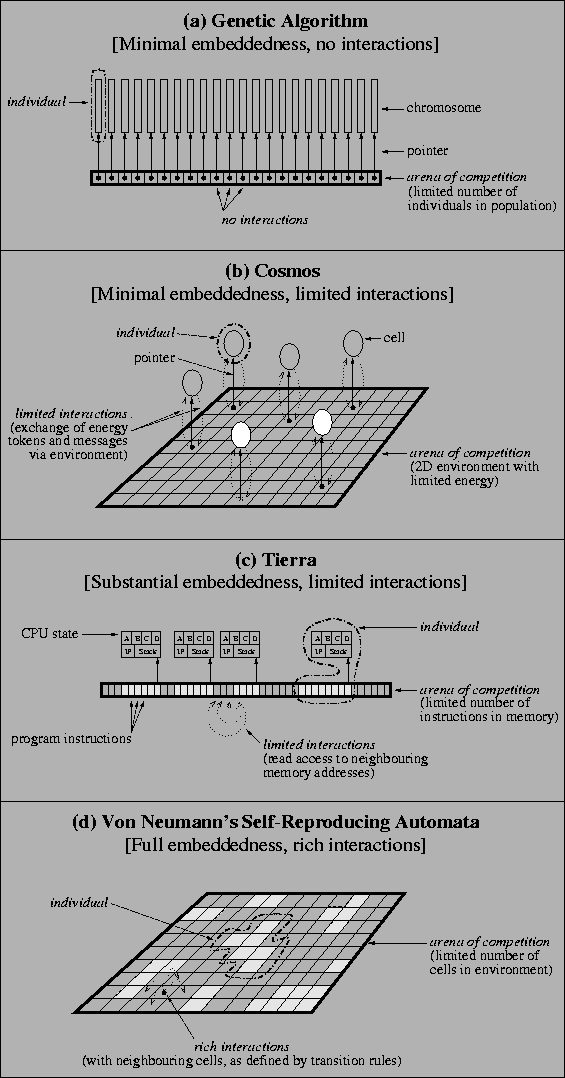 |
It should be emphasised that this notion of embeddedness is unrelated to the distinction between implicit and explicit encoding, which concerns the degree to which a process is governed by the environment (S in Definition 1) as opposed to a specific object situated within that environment (A in Definition 1). The notion of embeddedness exclusively concerns A rather than S, and, in particular, how much of A can be manipulated by other objects in the environment, and in what ways.
Related to the issue of physical embeddedness is that of how restricted is the range of interactions allowed between objects within the arena. In a standard GA, no direct interactions are allowed between chromosomes at all; the continued existence of an individual is decided by the externally-defined selection mechanism. In Cosmos, programs cannot directly interact with their neighbours, but they can exchange messages and energy tokens via the local environment. Although programs in Tierra are embedded in the arena of competition to a much greater extent than they are in Cosmos, the range of interactions allowed with neighbouring programs is still fairly restricted; programs can read the code of their neighbours, but they cannot directly write to neighbouring memory addresses.7.18 In contrast, von Neumann's cellular automata implementation is far less restrictive; the transition rules of the cellular automata define neighbourhood interactions which occur at the level of individual cells and which therefore do not respect boundaries between individual organisms.
From the point of view of the evolvability of individuals, the more embedded they are, and the less restricted the interactions are, then the more potential there is for the very structure of the individual to be modified. Sections of the individual which are not embedded in the arena of competition are `hard-wired' and likely to remain unchanged unless specific mechanisms are included to allow them to change (and the very fact that specific mechanisms are required suggests that they would still only be able to change in certain restricted ways). Additionally, recall from Section 7.1.2 that Pattee has argued that open-ended evolution fundamentally requires the evolution of new meaning in the system, and this can only be achieved in the context of a semantically closed organisation which is completely embedded within the physical world.
To end this section, I would like to briefly return to Holland's recent work with the Echo model ([Holland 95]: see Section 3.2.2). Echo possesses many of the features that I have just argued are desirable for a model of open-ended evolution. For example: selection in determined intrinsically by interactions between Echo organisms (or to use Holland's terminology, agents), rather than by an externally-defined fitness function; the process by which agents reproduce is implicitly defined in the Echo operating system rather than being explicitly encoded by individual agents; the agents are able to perform a variety of phenotypic behaviours; also, Echo is a material model, in which agents are composed of atomic units of matter and must collect raw materials from the environment before they are able to reproduce (I will say more about this topic in Sections 7.3.2 and 7.3.3). Echo is also designed upon more explicit design considerations than were most earlier artificial life models; the considerations for Echo are based upon a core set of principles which Holland believes are common to all complex adaptive systems. For all these reasons, I believe Echo represents a significant advance. However, the structure of the individual agents--the notion of what it is to be an agent--is still predefined, and the representation of agents is not fully embedded in the arena of competition. Additionally, the interpretation of agent's chromosomes is handled implicitly by the operating system. Now, the system was designed in this way because it is primarily intended as a general model of complex adaptive systems, rather than a specific model of biological evolution. Indeed, the various successful applications of Echo (mentioned in Section 3.2.2) testify to the value of the particular way in which the organism and environment structure have been predefined; if no higher-level structure were imposed, it would be difficult to model most complex adaptive systems of interest (e.g. ecologies, economies, etc.).
In the context of open-ended evolution, however, the design still has some shortcomings. The fact that the Echo operating system implicitly interprets the agents' chromosomes means that they can never come to encode anything more than the fixed range of actions (e.g. offence, defence, conditional exchange of resources) predefined by the designer. In Hidden Order, Holland discusses how new meaning can arise in a system, but acknowledges that Echo is deficient in this respect [Holland 95] (p.138). As Pattee has suggested ([Pattee 95b]: see Section 7.1.2), it is only when an organism's genotype, phenotype, and the interpretation machinery that produces the latter from the former, are all embedded in the arena of competition that fundamentally new symbolic information can arise in the genome (thereby permitting truly open-ended evolution). In the discussion of the desirable properties of proto-DNA in Section 7.2.3, it was suggested that this too would initially be interpreted implicitly. It was, however, stressed that the potential should exist for explicit interpretation machinery to evolve (although how this potential might be assured is an open question).