

Next: Self-Reproduction and Open-Ended Evolution
Up: Self-Reproduction and Evolution Revisited
Previous: Some Definitions
General Issues of Reproduction
The description of self-reproduction in the definitions of the
previous section is stated in terms of the configuration d of object
A
forcing surrounding
S
to produce a copy of
A.
It says
nothing of how exactly d can (or should) achieve this feat, although
this is a rather fundamental question when comparing different
sorts of reproduction. Indeed, when looking at any sort of
reproduction, I think it is useful to look at the process by which
reproduction is accomplished in (at least) three different
ways:
- 1.
- The degree to which the algorithm for reproduction (the way in
which the process is specified and controlled) is explicitly
encoded on the configuration being reproduced (cf. d in
Definition 1), rather than being implicit
in the physical laws of the world (cf.
S
in
Definition 1).
- 2.
- Whether reproduction happens purely by the action of the
physical laws of the world on the configuration to be reproduced
(auto-reproduction), or whether it also requires auxiliary
physical (or logical) machinery (assisted-reproduction). I
use the term auto-reproduction rather than self-reproduction here
because the latter is often used less specifically. I wish to
emphasise that this auto-assisted distinction is only one of a
variety of issues involved in the general concept of reproduction.
- 3.
- The number of different configurations that exist, connected by
mutational pathways, that are capable of reproducing their
specific form (i.e. the distinction between limited hereditary
reproducers and indefinite hereditary reproducers). From the
point of view of an individual reproducer, this can be
expressed in terms of the proportion of all possible mutations it
may experience that will result in the production of distinct, yet
viable, reproducers.
There are a number of points to note about these distinctions. First
it should be said that (3), in contrast to (1) and (2), does not
properly relate to individual reproducers per se, but rather to
lineages of reproducers. It is therefore not relevant when considering
self-reproduction in and of itself, but is an important factor when
considering the evolutionary potential of a class of reproducers.
Secondly, with regard to distinction (2) in the context of material as
opposed to logical systems, I do not consider the fact that objects in
material systems need to collect raw materials to be relevant.
As long as the surrounding
S
ordinarily contains sufficient raw
materials for a reproducing object
A
to build a copy of itself, and
that the configuration d of
A,
and the surrounding
S
between
them effect the collection of these materials to build the copy
without further assistance, then our definition of
self-reproduction given in Section 7.2.1 is still
satisfied.
The distinction between auto- and assisted-reproduction is a
dichotomy, but the other two distinctions each define a spectrum of
possibilities.
The distinctions are generally independent of each
other, although the more explicitly encoded the reproduction
algorithm is, the less likely, in general, it is to be an indefinite
hereditary reproducer (because of the increased chance of mutations
disrupting the copying process; see Section 7.2.3).
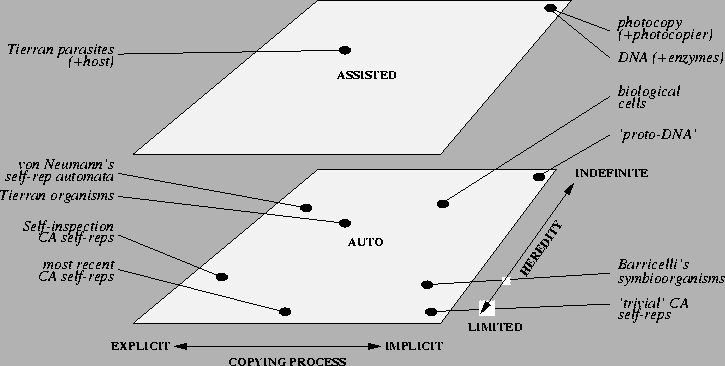
Figure 7.1 shows how some of the
reproducers that have been discussed so far can be categorised
according to each of these three distinctions. The diagram is not
supposed to be quantitatively accurate (not least because the
limited-indefinite heredity axis is in fact infinitely long, and also
because I have not offered any way of quantifying these factors), but
I have tried at least to highlight the general relationships between
different types of reproducers according to each of the three
distinctions.
Figure 7.1:
Categorisation of Reproducers.
|
|
There are a number of points about this diagram that require further
explanation:
- Tierran organisms and von Neumann's self-reproducing automata are
placed midway along the limited-indefinite hereditary scale
because, although both representations are capable of
supporting universal computation in principle, only mutations which
retain the ability to reproduce will be viable. In
particular, in von Neumann's architecture (described in
Section 3.2.1), a mutation which affects
a section of the tape which encodes the constructing automaton A,
the copying automaton B, or the control automaton C, will
generally disrupt the ability of the combined automaton to produce
viable offspring. Likewise, in Tierra, a mutation which affects a
section of the program which encodes the self-reproduction algorithm
will generally disrupt the ability of the program to reproduce. We
will return to these issues in Section 7.2.3.
- Trivial cellular automata self-reproducers (e.g. where the state of a single
cell is reproduced in neighbouring cells purely due to the CA's
transition rules) are, in general, limited
hereditary reproducers, because even though a single state may be able
to reproduce, a compound set of states will usually not be able to
reproduce as a whole.
Notice that in much of the recent artificial life work with
self-reproduction (e.g. [Langton 84] and other
studies mention in Section 3.2.1), the
distinction between trivial and non-trivial self-reproduction is
perceived to be a distinction on the implicit-explicit axis.
However, from an evolutionary point of view, the limited-indefinite
heredity axis is clearly the most relevant. Indeed, this is exactly
what von Neumann himself says:
``One of the difficulties in defining
what one means by self-reproduction is that certain organizations,
such as growing crystals, are self-reproductive by any naive
definition of self-reproduction, yet nobody is willing to award them
the distinction of being self-reproductive. A way around this
difficulty is to say that self-reproduction includes the ability to
undergo inheritable mutations as well as the ability to make another
organism like the original''
[von Neumann 49] (p.489).
Barry McMullin has presented an enlightening discussion on the history
of the confusion over von Neumann's work, which he refers to as the
`von Neumann Myth' (see, for example, Section 4.2.7 of
[McMullin 92a]). One result of this confusion has been that the
majority of subsequent research concerning this issue of trivial
self-reproduction has concentrated on the implicit-explicit
distinction, rather than the limited-indefinite heredity
distinction.7.8
- DNA reproduction is assisted, because it can only do so with the
aid of a host of enzymes to control the unwinding of the double
helical structure, the polymerisation of the individual bases of the
new molecule, etc. (see Section 7.2.3). At the same time,
DNA is, in the presence of suitable enzymes, capable of indefinite
heredity assisted-reproduction; the enzymes are able to copy any DNA
double helix, no matter what sequence of bases it comprises. Within
this context, the copying process is implicitly encoded in the DNA's
environment (in the enzymes which support the reproduction process,
and in the physical laws governing the inherent bonding affinities
of the bases) rather than being explicitly encoded upon individual
strands on DNA.
In contrast, a cell as a whole can be considered an
auto-reproducer, as it can completely direct its own reproduction
(in the presence of sufficient energy and raw materials from the
environment), but its hereditary potential is slightly more
restricted than DNA assisted-reproduction, because some mutations
may disrupt the ability of the cell to reproduce.
Similarly, each of Barricelli's symbioorganisms
(described in Section 3.2.2) can be
considered, collectively, as an auto-reproducer, although individual
digits within the symbioorganism are assisted-reproducers. The same
analysis can be applied to collectively-autocatalytic reaction
networks (e.g. [Kauffman 93]).
- Most importantly, I have placed the hypothetical `proto-DNA'
(i.e. a desirable seed for open-ended evolution) in the
auto-implicit-indefinite hereditary corner of the space. The seed
should be auto-reproducing (i.e. not rely upon auxiliary machinery)
if it is to have a reasonable chance of spontaneously emerging, and
it should be an indefinite hereditary reproducer to support an
on-going, open-ended evolutionary process. The requirement that it
be an indefinite hereditary reproducer is most easily fulfilled if
it reproduces implicitly. I will talk about this in more detail in
the next section.
Next: Self-Reproduction and Open-Ended Evolution
Up: Self-Reproduction and Evolution Revisited
Previous: Some Definitions
Tim Taylor
1999-05-29